Tous droits réservés © Ronald de Sousa
La biologie est la science du vivant. La philosophie, elle, s'interroge sur le sens de la vie, aussi bien que sur le sens des mots et des idées. On peut donc attendre de chacune de ces deux disciplines qu'elle mette l'autre à contribution. Dans ce livre, la philosophie invitera la biologie à nous dire qui nous sommes, en tant qu'organismes biologiques. La philosophie, de son côté, se chargera d'éclaircir le sens de certaines questions dont la simplicité n'est qu'illusoire : la vie a-t-elle un but? Comment la biologie serait-elle différente sur une autre planète? L'évolution est-elle une ascension vers des formes toujours supérieures? De quel genre de supériorité s'agirait-il? Y a-t-il des lois de la biologie? S'il y en a, doit-on s'attendre à ce que ces lois s'avèrent réductibles à celles de la chimie et de la physique? Ou est-ce que la science de la vie, au contraire, serait destinée à demeurer radicalement différente des autres sciences?
Nous commencerons par aborder, dans ce premier chapitre, la question la plus générale et peut-être la plus déroutante de toutes : qu'est-ce que la vie? Avant de faire l'inventaire des principales réponses possibles, nous aurons l'occasion de nous pencher sur certaines questions préalables concernant notre démarche philosophique elle-même : qu'est-ce qu'une définition? Y en a-t-il plusieurs sortes? Comment reconnaître celles qui sont adéquates?
QU'EST-CE QUE LA VIE?
L'ampleur de la question nous confronte un peu comme un immense mur lisse. On ne sait trop par où s'y prendre. Les questions typiques de la philosophie font souvent cet effet : qu'est-ce que le temps? le bonheur? l'intelligence? la beauté? Leur caractère insaisissable vaut sans doute à la philosophie une certaine mauvaise réputation : celle d'engendrer des débats aussi interminables que stériles, à propos de questions oiseuses et insolubles.
Nous reviendrons sur ce qui constitue à vrai dire une question philosophique. Mais commençons, pour ne pas donner prise trop vite à de telles accusations, par nous attaquer à un problème un peu plus maniable : comment distinguer, en pratique, ce qui est vivant de ce qui ne l'est pas?
La question s'est posée concrètement en 1996, lorsqu'on a fait beaucoup de cas d'une éventuelle découverte de traces de bactéries sur la planète Mars, ou plus précisément dans certains rochers tombés de la planète Mars il y a quelques milliers d'années. On se souviendra peut-être que le débat soulevé par cette découverte, d'abord fiévreux, s'est vite ramené à des questions extrêmement techniques au sujet de la forme exacte des isotopes de carbones faisant partie de certaines molécules. Débat pour le moins décevant pour qui veut savoir ce qu'est en vérité la vie, car il donne l'impression que la réponse n'est intelligible que pour certains spécialistes. Or il semble bien que la différence entre le vivant et ce qui ne l'est pas saute aux yeux, et qu'ele n'a guère à voir avec ces minuties scientifiques.
Par ailleurs, même du point de vue le plus rigoureusement scientifique, la question qui se posait au sujet des rochers de la planète Mars ne saurait être identique à celle qui nous occupe. Admettons que les spécialistes se soient mis d'accord que seule la présence de telle trace chimique indiquerait qu'il y eût des êtres vivants sur Mars. Il s'agirait là d'un indice, mais non d'une définition.
Pourquoi? La notion de définition exige un éclaircissement. Par ailleurs, le problème de la définition nous donnera l'occasion d'aborder une première fois certains des thèmes principaux qui nous occuperont dans ce livre. En effet, la nature de la définition prête à controverse en philosophie, et son rôle dans la biologie est particulièrement problématique.
LA DÉFINITION
On entend souvent dire que la rigueur scientifique et philosophique exige la définition des termes utilisés. Mais qu'est-ce, au juste, que définir un terme?
Pour définir la définition, il convient d'abord d'en distinguer deux sortes. On pourrait les baptiser définition de dictionnaire et définition encyclopédique. Ces appellations ne sont que des aide-mémoire, car on trouve souvent dans les dictionnaires des renseignements d'ordre encyclopédique, et inversément. Cependant les dictionnaires visent généralement à fournir des expressions synonymes du mot à définir, alors que les encyclopédies aspirent plutôt à nous renseigner sur la nature de la chose. Par conséquent, il suffit de connaître la langue pour être en mesure de déterminer la justesse d'une définition de dictionnaire. Cela n'est nullement suffisant, par contre, pour trancher de la valeur d'une définition de type encyclopédique. Lorsqu'on approfondit une définition à l'aide de données scientifiques, on peut arriver à des formules que le sens commun n'aurait eu aucun moyen d'entrevoir. Témoin, par exemple, la définition scientifique de l'eau : « H2O » précise sa composition chimique, et donc présuppose toute une théorie de la matière chimique, vieille de moins de trois siècles. Elle ne se laisse évidemment pas deviner sur la base du sens familier d'un mot dont chacun sait depuis toujours qu'il désigne un liquide incolore, inodore, et potable.
On verra sous peu qu'il y a des cas difficiles à classer, et que certains de ces cas limites sont précisément ceux qui intéressent la biologie. Les deux types ont pourtant ceci en commun : toute définition vise à préciser ce qui est inclus par un terme donné et ce qui en est exclus. Une bonne définition opère un triage bien défini entre ce qui rentre et ne rentre pas dans son acception : elle précise des propriétés nécessaires, c'est à dire dont le manque garantira l'exclusion de la chose en questions, et (s'il y a lieu) une ou plusieurs propriétés suffisantes pour qu'elle soit incluse. Ainsi, pour répondre au terme de « mère », il est nécessaire d'avoir un enfant; mais cette condition n'est pas suffisante, puisqu'il faut aussi remplir une seconde condition nécessaire, celle d'être femelle.
Il importe de distinguer aussi deux modes d'inclusion ou d'exclusion. Le premier se nomme intensionnel. Il s'agit de l'inclusion d'une notion dans une autre. Le second, appelé extensionnel, porte sur la classe des individus désignés par la notion en question. Cette distinction se comprendra plus facilement à partir d'un exemple. Pour ne pas trop nous éloigner de la biologie, prenons le terme de mammifère, que le Petit Robert définit comme suit :
S'agit-il là d'une définition de dictionnaire ou encyclopédique?
Hélas, le cas n'est justement pas clair. Certes, toute personne éduquée qui sait le français sera plus ou moins en mesure d'entériner la définition du Petit Robert. Mais on imagine facilement des cas où l'on puisse se tromper sur la suffisance ou la nécessité de certains traits. Les enfants sont parfois surpris d'apprendre, par exemple, qu'il y a des mammifères volants et des mammifères aquatiques. La définition n'en dit rien : elle ne les exclut donc pas. Mais jusqu'au moment où l'on apprend que les chauves-souris ne sont pas des oiseaux, et que les baleines et les dauphins ne sont pas des poissons, rien n'empêche de penser que la définition des mammifère exclut implicitement les habitants de l'air et de l'eau.
Il s'agit donc ici d'une définition qui exprime à la fois le sens du terme, tel qu'il est compris par ceux qui savent l'utiliser, et la nature de la chose en question, telle qu'elle a été découverte par la recherche scientifique. Elle est donc à la fois de dictionnaire et encyclopédique. Voilà qui est typique des définition d'ordre scientifique. En effet, l'élaboration d'une telle définition repose d'abord sur certaines intuitions concernant le sens des mots en question. Sans doute, pour beaucoup, l'extension de « mammifère » a commencé par exclure ceux qui ressemblaient superficiellement à des poissons, ou à des oiseaux. Ce n'est que grâce à des observations plus attentives qu'on a pu remarquer que certains animaux équipés d'ailes ou de nageoires jouissaient de ressemblances plus profondes et plus générales avec les mammifères les plus typiques qu'avec les poissons ou les oiseaux. Plus tard encore, comme nous aurons l'occasion de le voir à loisir au chapitre six, les techniques d'analyse de la matière génétique ont confirmé et précisé ces jugements de ressemblance. Grâce à elles, il nous est maintenant permis d'affirmer qu'il ne s'agit pas simplement de ressemblances, mais véritablement de rapports de parenté. Ainsi, il est littéralement vrai que les loups, les humains, les chauves-souris et les baleines ont des ancêtres en commun que ne partagent ni les poissons ni les oiseaux. Il s'agit là donc très clairement de définitions encyclopédiques dans le plein sens du terme, puisqu'aucune réflexion sur le sens des mots n'aurait pu amener cette conclusion.
Une définition intéressante de la vie sera donc, de toute évidence, une définition encyclopédique. On peut cependant se mettre à sa recherche en s'appuyant sur les intuitions que nous dicte le sens commun, tout en rappelant que ce point de départ ne garantit nullement que l'aboutissement de l'enquête soit, elle aussi, évidente à la seule lumière du bon sens.
Les intuitions du sens commun portent sur les deux modes d'inclusion dont il a été question, c'est à dire à la fois sur le sens du mot et sur son extension. Sur le mode intensionnel, on dira que le sens même du mot vie comprend certaines capacités, sans lesquelles rien ne serait vivant. Le mode extensionnel, rappelons-le, se rapporte aux choses : il s'agira donc de déterminer quels individus sont à ranger dans la catégorie des êtres vivants. Ces deux sources d'intuition donnent parfois des résultats contradictoires. Leur confrontation peut alors mener, par voie d'une sorte de dialectique, à un raffinement des définitions dont on était parti.
Voyons comment pourrait se dérouler une telle dialectique, à partir de ce que nous dicte le sens commun sur le sens du mot vie et sur son extension.
SIX RÉPONSES POSSIBLES
Ce qui frappe tout d'abord chez les êtres vivants, c'est qu'ils bougent tout seuls. (Ainsi Xénophane, philosophe grec du 6ème siècle avant notre ère, avait proclamé que les roches aimantées avaient une âme.) Cette constatation nous suggère une première définition :
Déf. 1 : Le vivant est ce qui jouit de motricité autonome. Cette définition est-elle adéquate? La question peut prêter à controverse. On peut s'en rendre compte en imaginant un petit dialogue entre un défenseur, D, et un opposant, O, de cette définition. Ce dialogue fera ressortir le problème qui menace toute tentative de définition en philosophie, c'est à dire le caractère discutable des termes qui entrent dans la définition elle-même.
O : Dans ce cas, il faut rejeter cette définition. Car, de toute évidence, certains organismes vivants ne bougent pas : c'est plus ou moins ce qui distingue les plantes des animaux. Bouger tout seul ne constitue donc pas une condition nécessaire. [Notons que cet argument fait lui aussi appel au sens commun, puisque il traite comme évident le fait que les plantes sont des êtres vivants : il s'appuie, autrement dit, sur un parti pris concernant l'extension du terme « vivant ».]
D : Mais si, les plantes bougent bel et bien toutes seules. Mais elles bougent lentement et par croissance, non pas, à quelques exceptions près, par locomotion.
O : Soit : ça dépend en effet de ce qu'on veut dire par bouger. Mettons peut-être que la condition soit nécessaire. En tous cas, elle n'est pas suffisante. Sinon, un glacier, qui lui aussi bouge lentement tout seul, ou même les planètes, seraient des êtres vivants.
D : Mais le mouvement des planètes et des glaciers n'est pas de la bonne sorte! Ces mouvements s'expliquent entièrement à partir des forces auxquelles planètes et glaciers sont assujettis selon les lois de la nature. C'est donc qu'ils ne bougent pas à proprement parler tout seuls, comme l'exige notre définition.
O : Est-ce à dire que la différence entre les glaciers ou les planètes et les êtres vivants est que ces derniers échappent aux lois de la nature? Voilà qui serait paradoxal....
D : Non, mais...
O : Laissons donc de côté les glaciers et les lois de la nature. On conçoit cependant que certains objets -- des robots, peut-être -- bougent tout seuls, sans pour autant qu'ils soient vivants. [Ici encore, l'argumentation fait appel au sens commun, qui juge exclut les robots de l'extension du terme « vivant ».))
D : Notre difficulté vient du fait que l'expression « bouger tout seul » n'est pas claire. Si on la comprend correctement, on s'apercevra que tous les êtres vivants, et rien qu'eux, bougent tout seuls. Les robots, eux, ne bougent pas vraiment tout seuls puisqu'ils ne marchent plus dès que leur pile électrique est déchargée....
O : Mais nous non plus, nous ne bougeons plus si on nous coupe nos sources d'énergie! Bouger tout seul ne saurait vouloir dire : bouger sans utiliser de source externe d'énergie. Car justement tout ce qui vit sur terre a besoin de l'énergie qui nous vient du soleil -- soit directement, comme c'est le cas de la photosynthèse chez les plantes, soit indirectement, comme c'est le cas des animaux qui digèrent la nourriture qu'ils absorbent.
D : Sans doute; mais l'être vivant absorbe et utilise l'énergie de manière naturelle, à la différence du robot. C'est ce qui fait qu'il bouge vraiment tout seul dans le sens qui nous intéresse.
O : Mais ce « sens qui t'intéresse », de toute évidence, n'est autre qu'un sens tout à fait arbitraire qui te permet d'insister sur ta définition! Te voilà donc à tourner en rond : en effet l'expression « bouger tout seul », une fois affublée du qualificatif « naturel », n'a plus son sens habituel. Il est donc impossible de vérifier si oui ou non cette condition est en effet nécessaire et suffisante pour marquer la différence des êtres vivants....
Commençons par une tentative de rafistolage, visant à remédier aux défectuosités de la définition que nous venons de débattre. Dans le débat auquel nous venons d'assister, l'exemple du glacier n'était assurément guère convaincant. Les chances du robot semblaient meilleures. Or, ce qui différencie le robot du glacier, c'est que le robot forme une unité organique qui semble rivaliser avec celle des êtres vivants. Cette observation suggère une deuxième définition possible.
Déf. 2 : Le vivant est ce qui possède une intégrité organique individuelle. Cette définition ne sonne pas trop mal. Elle illustre pourtant une nouvelle difficulté assez caractéristique de toute définition qui se veut tant soit peu substantielle, à savoir que les termes de la définition elle-même sont peu utiles, soit parce qu'ils sont difficiles à comprendre, soit parce qu'ils se réfèrent eux-mêmes implicitement au terme que l'on cherche à définir. En effet, qu'est-ce que l'intégrité organique, si ce n'est justement le rapport qui unit les organes d'un être vivant?
Cette formule nous permet pourtant de faire un pas en avant. Elle propose au moins un critère solide, précisément en ce qu'elle présuppose que tout être vivant est un ensemble d'organes, c'est à dire de parties qui ont des rapports coopératifs entre elles. Il ne s'agit pas simplement d'un amoncellement quelconque.
Cette caractéristique range bien le mouvement de l'animal ou du robot d'une part, et celui du glacier de l'autre. Par contre, si l'on veut écarter les robots -- du moins provisoirement -- la définition 2 ne nous en donne pas les moyens. Car les différentes parties d'un robot, leurs « organes », sont bien certainement liés entre elles par des relations coopératives, d'autant plus claires qu'elles ont été établies exprès par les constructeurs du robot. Toute partie dont la présence ne s'explique pas par sa contribution au tout, sera par là condamnée à être supprimée dans un modèle perfectionné. C'est chez les êtres vivants, au contraire, qu'il semble subsister des « organes » qui n'en sont pas -- des parties du corps, appendice, amygdales, ou mamelles d'homme, qui ne servent apparemment à rien. Faut-il conclure qu'un robot est plus vivant qu'un animal? Évidemment non. C'est qu'ici encore, nous sommes assez sûrs de l'extension du terme pour récuser toute définition qui entraînerait cette conséquence paradoxale. C'est la définition qu'il faut encore modifier.
Par ailleurs, l'intégrité de l'individu pourrait se comprendre par rapport à ce qui est borné par la frontière de sa peau. Un individu comme nous le concevons typiquement occupera un certain espace. La peau sépare l'intérieur de l'extérieur. Mais la peau n'est pas qu'une simple limite : elle est elle-même un organe. C'est dire qu'elle sert les intérêts de ce qui est à l'intérieur au dépens, s'il y a lieu, de ce qui est à l'extérieur.
Deux précisions nécessaires compliquent toutefois la notion simple d'une peau qui ne serait que la frontière entre l'individu et le milieu environnant.
1. La peau est un organe qui fait partie de l'organisme : ce n'est donc pas la frontière entre les l'organisme et le milieu. Elle est plutôt comparable à une zone de transition, qui de par l'activité constante des pores d'une part et sa continuité avec les muqueuses de l'autre, mène un trafic constant entre l'extérieur et l'intérieur. On peut tout de même parler d'extérieur et d'intérieur, puisque ce qui passe cette zone devient par là aliment -- nourriture ou oxygène -- lorsqu'elle vient de l'extérieur, ou déchet, lorsqu'elle passe dans l'autre sens. Mais rien n'est aliment ni déchet, si ce n'est en vertu de ce passage, puisque ces termes n'ont de signification que par rapport aux rôles que jouent les substances en question dans la vie de l'organisme.
2. Il peut y avoir une interdépendance étroite entre le milieu et ce qui est en decà de la peau. En effet, si tout individu semble limité par son épiderme, les actes qui le lient au monde sont contrôlés tout autant par les circonstances dites extérieures que par les systèmes de contrôle (système nerveux ou hormonal) qui font partie de l'organisme physique. Cela est clair dans le cas des tropismes (1) qui régissent, chez les organismes les plus simples, un comportement qui dépend entièrement d'une source de lumière ou d'un gradient chimique. Mais on peut aussi bien le constater dans le cas d'une simple action, comme celle de tendre la main afin de saisir une tasse de café. Les contractions musculaires voulues sont, bien entendu, modulées par le cerveau. Mais comment se fait-il que ma main ralentisse exactement au bon moment pour que l'anse de la tasse soit à portée de mes doigts, sans la heurter ni la dépasser? C'est que tout ce processus met en jeu un circuit cybernétique dont fait partie non seulement mon cerveau et mon appareil visuel, mais aussi la tasse elle-même. La position de celle-ci opère donc un téléguidage, dont le processus passe par mon cerveau mais n'est nullement confiné à l'intérieur de ma peau. L'intégration fonctionnelle des organes, c'est à dire leur coopération, ne se passe pas sans la participation d'un milieu qui n'est pas, lui, un organe.
Peut-être peut-on espérer éclaircir la distinction entre organe et environnement en précisant plus exactement la notion de coopération dont il s'agit. Co-opérer, c'est littéralement travaillerensemble; et travailler ensemble, c'est avoir un certain but en commun, en fonction duquel s'expliquent les activités des « organes » en question. Mais quel but, fixé par qui ou par quoi? Si l'on peut dire que le robot a un but, c'est que justement les ingénieurs l'ont conçu et construit pour qu'il soit utile à quelque chose. Le but d'un robot ne réside pas en lui-même, mais dans son utilisateur ou dans son créateur.
Voilà qui nous ramène à un aspect supplémentaire de la notion d'intégrité individuelle. S'il s'agit vraiment d'un individu, et non pas simplement d'une région dans l'espace hétérogène arbitrairement délimitée, alors on doit pouvoir faire la différence entre ce qui favorise la perpétuation de cet individu et ce qui peut lui nuire. C'est à l'individu que servent le plus directement ses organes, puisque le fonctionnement de ceux-ci est en général indispensable à l'existence de celui-là.
Cette constatation incite à formuler une troisième tentative de définition.
Déf. 3 : Le vivant est ce qui possède une finalité intrinsèque, c'est-à-dire d'un but qui appartient à l'organisme lui-même et non à quelque autre être dont il est l'instrument. Cette définition rend bien compte de l'idée que tout individu vivant est équipé d'organes dont la fonction est de pourvoir à sa survie. Cependant, si la finalité d'un
organe s'explique relativement facilement par rapport à l'organisme dont il fait partie, celle de l'organisme entier n'est pas du tout si claire. A quoi sert un homme? A quoi sert un chat? A quoi sert une fleur? La tradition philosophique et surtout théologique n'a certes pas été à court de réponses à ce genre de questions. Mais ces réponses ne réussissent guère à apaiser notre perplexité.
Il y a d'une part une certain tradition théologique, qui veut que nous soyons les instruments des desseins du Créateur. Nous serions alors, en quelque sorte, les robots du bon Dieu. Idée pittoresque, sans doute, mais dont la véracité ne serait nullement suffisante pour démontrer que nous sommes ni plus ni moins vivants que des machines. D'autre part, une certaine conception biologique de l'évolution voudrait que toute vie individuelle n'ait pour but que la reproduction de son patrimoine génétique. Dans cette optique, toute finalité intrinsèque qui semblerait régir un être vivant ne serait qu'illusion. Cependant, il est clair qu'un tel refus de toute finalité individuelle, quel que fût son bien-fondé, ne suffirait pas non plus à invalider le statut d'être vivant qui est l'apanage des bactéries, des plantes, et des animaux.
L'idée de finalité, et le rôle capital qu'elle semble jouer dans la biologie, nous occupera beaucoup dans la suite de ce livre. (Voir surtout les chapitres 2, 4 et 7). Nous nous bornerons, pour le moment, à souligner la difficulté, voire l'impossibilité, de juger en pratique du bien-fondé de l'attribution d'un but ou d'une finalité à une entité dont aucun dessinateur n'a dressé les plans ni précisé la raison d'être. De fait, l'hypothèse théologique d'une intelligence créatrice qui fît tout pour le mieux dans le meilleur des mondes venait bien à propos combler une lacune scientifique. Elle permettait de postuler l'existence d'une finalité spécifique à chaque espèce vivante (sinon de toujours en deviner la nature). On pouvait alors, sans trop forcer et ne fût-ce que par ricochet, accorder une sorte de finalité intrinsèque à tout individu qui pût se prévaloir d'appartenir à une espèce façonnée dans un but précis par le Créateur. Or, depuis Laplace, voire depuis Lucrèce, la science se targue de ne pas avoir besoin de cette hypothèse. C'est se rendre la tâche sensiblement moins facile; mais la philosophie, encore moins que la science, ne peut s'y soustraire. L'idée de finalité, et à plus forte raison l'idée d'une finalité intrinsèque, demeue trop opaque pour que nous puissions en tirer un critère valable. Le prochain chapitre tâchera de démêler les questions enchevêtrées qu'elle soulève. Mais pour le moment, laissons de côté les critères découlant de la finalité.
Le reste de ce chapitre sera consacré à trois autres critères visant à marquer la différence du vivant. Les deux premiers se présentent comme des définitions de dictionnaire. Le dernier, par contre, relève de véritables découvertes scientifiques. Cette dernière tentative de définition ne sera pas plus évidente au yeux du sens commun que l'est la définition chimique de l'eau, H2O. Elle nous ouvrira cependant une perspective surprenante, susceptible d'éclairer sous un jour radicalement neuf la nature profonde de la vie.
La quatrième et la cinquième définitions ont trait à deux caractéristiques dont la possession chez les êtres vivants typiques ne fait aucun doute : la capacité d'assimiler des substances nutritives tirées du milieu environnant (métabolisme), et la capacité de produire de nouveaux êtres construits plus ou moins suivant le même modèle (reproduction). Si nous prenons ces capacités une à une, c'est qu'il faudra s'interroger sur leur indépendance : l'une pourrait-elle exister sans l'autre?
Déf. 4 : Le vivant est ce qui est capable de métabolisme, c'est-à-dire de reconstruire sa propre substance organique à partir de l'énergie et de molécules ou d'ions chimiques tirés du milieu environnant. La capacité des êtres vivants à cet égard est banalisée quotidiennement par les animaux comestibles. En effet, ceux-ci sont de véritables usines à transformer l'herbe en chair. Les plantes, elles, opèrent une transformation de la matière un peu moins spectaculaire, puisqu'elles tirent du sol et de l'air les matières premières dont elles se constituent et les assimilent sans être obligées de les extraire au préalable de composés chimiquement plus complexes. Mais ce n'est là qu'un détail, puisque, dans tous les cas, il se passe une transformation chimique qui intègre un matériau tiré de l'extérieur à la substance organique de l'individu.
Ce fait fondamental rejoint, tout en les éclaircissant, les deux caractéristiques précédentes. Dans un sens très concret, le métabolisme entretient l'intégrité organique de l'individu, puisque c'est par lui qu'est franchie la barrière qui sépare l'individu de son environnement. Et d'autre part, dans un sens tout aussi concret, on peut constater que le métabolisme sert littéralement la finalité de l'individu, puisqu'il lui permet de remplacer sa substance et de survivre en tant qu'organisme distinct.
Cette quatrième définition semble donc prometteuse. Il faut pourtant émettre deux réserves, qui mettent en doute non pas la suffisance, mais la nécessité de la capacité métabolique. La première réserve soulève une exception possible; la deuxième découle de certaines spéculations qui tiennent de la science-fiction, ou plutôt de la recherche sur la vie artificielle. Commençons par celle-ci.
Nous verrons plus tard en plus de détail quels sont les problèmes soulevés par la tentative de créer une vie artificielle (chapitre 9). Il ne sera pas question des techniques de reproduction artificielle. Ce dernier terme recouvre la micro-injection directe d'un spermatozoïde (la cellule gamétique portant le patrimoine génétique du mâle) dans le cytoplasme de l'ovocyte (la cellule gamétique femelle); la fécondation in vitro (« en éprouvette ») qui est suivie de l'introduction de l'embryon dans la cavité utérine; le clonage, ou multiplication d'une cellule issue d'un animal adulte (un mouton anglais, « Dolly », produit par cette technique fit beaucoup couler d'encre en 1997), etc. Toutes ces techniques interviennent dans la reproduction biologique, et sont donc dans un certain sens des mesures « artificielles ». Mais elles ne prétendent pas manufacturer une forme de vie non biologique.
Or, certains informaticiens ambitionnent de créer une sorte d'être vivant non biologique, au même titre que d'autres travaillent à façonner une intelligence artificielle qui s'incarneraient non pas dans la matière organique mais dans le silicone ou autre matière inorganique. Une telle entreprise est-elle possible? Il semble bien qu'on ne puisse l'exclure à priori; d'ailleurs, la science-fiction nous habitue déjà à considérer l'idée comme banale. Banale ou non, elle est pourtant exclue par la définition trop étroite que nous venons de proposer. En effet, à supposer qu'une forme de vie non-organique puisse exister, on ne pourrait l'enfermer dans une formule qui se rapporte à l'assimilation de molécules ou d'ions chimiques dans une substance organique.
Néanmoins il semble que l'on puisse définir un équivalent plus abstrait du métabolisme. Ce serait l'idée, proposée par certains chercheurs contemporains, d'autopoïèse.
Le terme vient de deux mots grecs auto et poïesis, qui signifient respectivement soi-même et production; il connote donc un objet qui a la faculté de se créer lui-même, de maintenir sa propre structure et de se régénérer. L'autopoïèse caractérise donc toute unité constituée par une organisation de parties plus ou moins constante malgré les changements qui peuvent intervenir dans l'environnement. Non seulement son organisation interne contribue à maintenir l'unité de l'ensemble, mais aussi elle a la capacité de recréer sa propre structure dynamique, en dépit de l'influence de circonstances externes qui auraient tendance à entraîner sa désagrégation (à condition, bien sûr, que le pouvoir destructif de ces circonstances externes ne dépasse pas certaines marges de tolérance). On voit qu'il s'agit là d'une notion plus abstraite que le métabolisme, puisqu'il n'est précisé aucune limitation quant au type de matière qui pourrait le réaliser. Cependant le métabolisme peut être considéré comme un cas particulier de l'autopoïèse, puisque les systèmes digestifs et respiratoires (qui sont les piliers principaux du métabolisme) remplissent précisément les fonctions que nous venons d'attribuer à l'autopoïèse. En effet, on peut constater que les organes des êtres vivants, grâce à leurs fonctions métaboliques, remplissent parfaitement la condition ci-dessus: ils contribuent à maintenir l'unité de l'ensemble, et ont la capacité de recréer leur propre structure dynamique, en dépit de l'influence de circonstances externes qui auraient tendance à entraîner leur désagrégation.
L'avantage de la notion d'autopoïèse réside justement dans sa plus grande abstraction. Elle n'exclut pas nécessairement la possibilité d'une vie qui ne soit pas organique, tout en étant entièrement compatible avec la vie telle que nous la connaissons.
L'autopoïèse n'enraye pas, cependant, le premier motif de doute auquel nous venons de faire allusion. Ce motif fait valoir un contre-exemple. En effet, il y a un type d'entité qui se situe en marge des êtres vivants proprement dits, et que certains veulent inclure, alors que d'autres l'excluent de cette catégorie. Je veux parler des virus.
Les virus n'ont pas la capacité d'autopoïèse : ils sont entièrement dépourvus d'organes de métabolisme. Un virus consiste essentiellement en un tronçon cohérent de substance génétique, qui contient les instructions nécessaires à sa propre duplication. Mais étant dépourvu du cytoplasme qui permet à la moindre bactérie de maintenir sa structure, il n'a aucun moyen propre d'assimiler l'énergie ni les éléments du milieu environnant en sa propre substance. Il doit emprunter au milieu les ressources matérielles et énergétiques dont il a besoin pour se reproduire. Par conséquent, si on se tient à la définition que nous venons d'exposer, les virus doivent être exclus du domaine du vivant puisque cette condition nécessaire leur fait défaut.
Il y a cependant de bonnes raisons de classer les virus parmi les êtres vivants. Chaque virus a une structure caractéristique, qui suffit peut-être à définir son intégrité individuelle. Suivant la deuxième définition proposée plus haut, voilà qui suffirait à le qualifier d'être vivant. De plus, il semble que l'on puisse parler (ne fût-ce qu'au sens large) du comportement d'un virus, et que ce comportement puisse s'expliquer à partir d'une finalité propre : le virus satisferait donc également à la troisième définition. De fait, cette finalité semble n'être rien d'autre que sa propre reproduction. Et sa capacité de reproduction, même dépourvue de toute capacité de métabolisme, peut sembler suffisante pour en dicter l'inclusion dans le domaine du vivant. C'est que la reproduction à elle seule peut paraître un critère de vie à la fois nécessaire et suffisant. Voici donc la cinquième définition :
Déf. 4 : Le vivant est ce qui est capable de se reproduire. La notion de reproduction appelle quelques commentaires. On l'applique libéralement tant aux bactéries qu'aux plantes ou aux animaux. Or, dans le sens littéral, il n'y a que les organismes unicellulaires, ainsi que certaines plantes, qui se reproduisent véritablement. Lorsqu'on parle de reproduction sexuée, on entend le terme dans une acception élargie, puisque la reproduction sexuée ne reproduit aucun organisme existant. On devrait plutôt parler de production sexuée. En effet, rappelons que la soi-disant « reproduction » sexuée engendre une combinaison chaque fois absolument nouvelle d'éléments génétiques provenant (dans le cas le plus typique) d'une sorte de tirage au sort. La loterie génétique donne à chaque élément, c'est à dire à chaque gène, cinquante pour cent de chances d'être impliqué dans la nouvelle formule. On peut calculer que chez un animal comme l'être humain, qui possède 23 chromosomes, par exemple, le nombre de zygotes différents qui pourraient résulter de l'union de deux gamètes (oeufs ou spermatozoïdes) génétiquement distincts dépasse de loin le nombre d'atomes dans l'univers entier (2). C'est donc dire que dans le nombre démesuré des individus vivants appartenant à des espèces qui se servent de la reproduction sexuée (c'est-à-dire 95% de toutes les espèces), il n'y a pratiquement que des formes absolument uniques.
Cette variété ahurissante des formes vivantes mériterait peut-être elle-même d'être mise en vedette comme essentielle à la vie. Le problème est qu'il est difficile, en dehors du monde biologique, de quantifier objectivement la variété. (En chimie, les éléments se comptent par centaines; les corps composés par millions. Mais comment s'y prendre pour compter les espèces de choses?) Nous aurons l'occasion d'examiner plus loin (au chapitre 8) les conséquences de cet appareil de production de formes vivantes nouvelles qu'est la reproduction sexuée. (Nous continuerons, en dépit des réserves qui viennent d'être formulées, à nous servir du terme de « reproduction » consacré par l'usage.) Il suffira, pour le moment, de signaler que la reproduction au sens littéral, c'est à dire la division cellulaire qui amène de simples copies conformes de la cellule originale, perpétue l'existence non pas d'un objet particulier, mais d'un certain type d'objet.
La distinction qui vient de se profiler est importante. Il s'agit de la différence entre un objet particulier, qu'on peut désigner par le terme d'exemplaire, et un type d'objet. Cette différence prendra une importance capitale par la suite. En voici une illustration :
Combien y a-t-il de mots entre crochets dans la ligne suivante?
[LE MOT « MOT » DIFFERE DU MOT « MOT »]
Si l'on répond Sept, c'est que l'on a compté les exemplaires de mots. Si l'on compte les types de mots, par contre, il n'y en a que quatre : LE, MOT, DIFFERE, et DU.
Lorsqu'on parle de l'identité d'un élément linguistique, on peut vouloir parler soit du type, soit de l'exemplaire. La même ambiguïté peut se présenter dans un contexte biologique. Penchons-nous sur la paramécie, amibe unicellulaire qui constitue déjà un organisme complexe, muni de fonctions métaboliques. La cellule ingère des aliments qui lui sont nécessaire, les assimile à son propre cytoplasme, l'utilise pour remplacer ses organelles et ainsi de suite -- bref, elle transforme ce qui est étranger en sa propre substance.
Ce processus métabolique maintient en vie cet exemplaire particulier de paramécie. Cela contraste en apparence avec ce qui se passe dans le cas de la reproduction du même organisme, qui se fait par mitose ou division cellulaire. En effet, celle-ci donne lieu à deux exemplaires différents du même type.
Toutefois, pour deux raisons, ce contraste entre l'entretien d'un seul et même individu et la production d'individus nouveaux est peut-être moins important qu'on ne le croit.
En effet, si l'on parle de l'identité d'un organisme à travers le temps, il ne s'agit pas en fait de la notion la plus stricte d'identité. Celle-là exige que si a=b, toutes les caractéristiques de a appartiennent aussi à b. Or les caractéristiques, voire les éléments constitutifs, d'un organisme à un moment donné peuvent être largement différents de ceux qui le distinguent à un autre moment. Si donc on dit que la paramécie individuelle -- l'exemplaire particulier -- subsiste à travers le temps, c'est en dépit du fait que sa matière change constamment. Il s'agit donc d'un type d'identité qui n'est pas la plus stricte, mais qui tient, d'une part, de la constance de la structure autopoïétique qui effectue le processus d'assimilation de la matière étrangère, et, d'autre part, du fait qu'on peut suivre un tracé continu du lieu occupé par une telle structure dans le temps et l'espace.
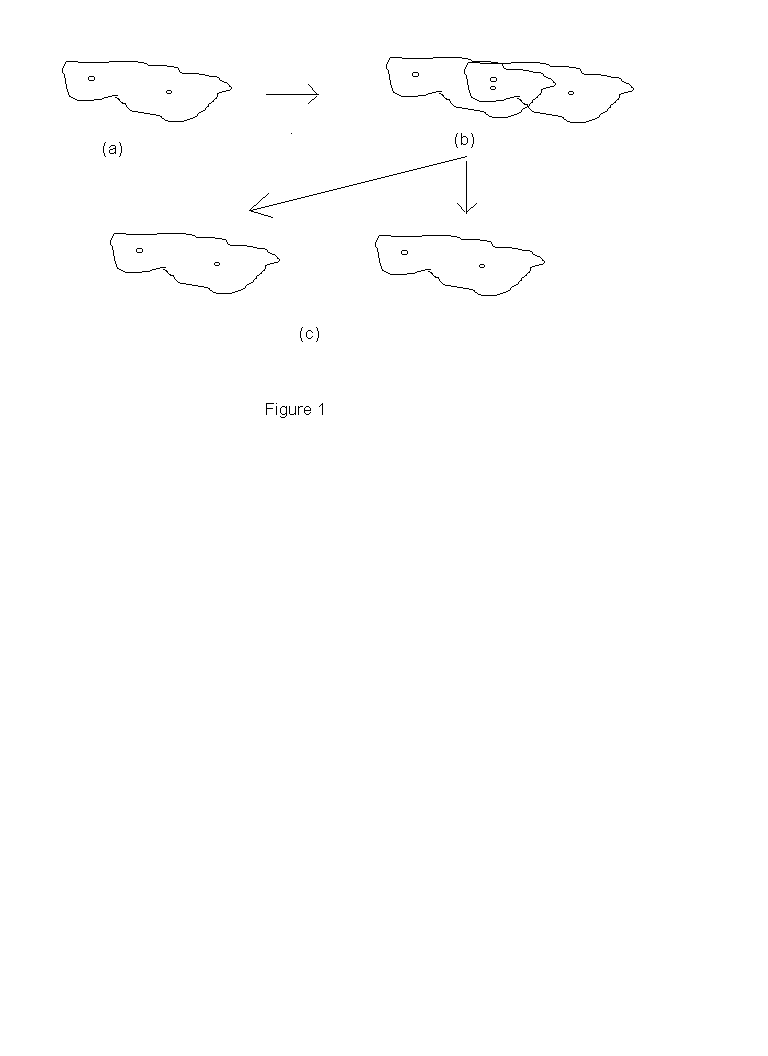
Or, on peut constater qu'en réalité la mitose n'implique pas, elle non plus, de discontinuité entre la paramécie « mère » et ses deux cellules « filles ». Il est vrai qu'il y a discontinuité spatiale entre les deux cellules filles; mais dans une perspective spatio-temporelle, non moins légitime, on peut parler d'un organisme qui trace une forme de Y, dont les deux cellules-filles sont les branches. Ainsi les étapes successives d'un processus de fission cellulaire (Figure 1(a)-(c)) peuvent être considérées comme des tranches instantanées d'un seul organisme à quatre dimensions (Figure 2 (a)-(c).)
Ces constatations impliquent la conséquence que voici : ce qui reste constant à travers les changements de matière entraînés par le métabolisme, c'est la structure de la cellule. Or, celle-ci relève non pas de l'exemplaire mais bien du type. En effet, au sens le plus strict, il ne peut y avoir identité d'exemplaire particulier là où il n'y a pas d'identité de matière.
La leçon à tirer de cet exemple est que, dans la perspective de l'espace-temps à quatre dimensions, le métabolisme, non moins que la reproduction, préserve non pas les exemplaires individuels, mais bien les types. Le contraste entre les deux phénomènes se limite au fait que seule la reproduction peut avoir pour résultat un nombre plus élevé d'exemplaires d'un même type. En vérité, la reproduction et l'autopoïèse peuvent être considérées comme deux stratégies visant le même but, c'est-à-dire la perpétuation de certains types de structures.
Dans cette optique, l'anomalie que présente le cas du virus est moins nette. Sa « finalité » est effectivement de perpétuer son type, qui se trouve être un arrangement caractéristique d'ADN(3).
Ces considérations suggèrent une conclusion plus spéculative et plus controversable : l'essence de tout être vivant ne tient pas à son identité particulière, mais bien au type qu'il représente. Ces deux catégories, exemplaire et type, se confondent facilement en pratique. La raison en est que dans la majorité des espèces -- les organismes sexués -- il n'y a généralement qu'un seul exemplaire individuel par type.
Or la notion d'un type est une notion informationnelle. Le sens spécial donné ici à cette expression provient en grande partie par la constatation que nous venons de faire : si ce qui fait l'identité d'une chose comprend sa matière propre, le remplacement de cette matière en fera une chose numériquement différente (4). Par contre, si l'on accepte comme équivalente à une chose une autre du même type, cette notion est informationnelle, puisqu'il suffit, pour établir cette identité, que l'une soit conforme à l'information qui serait contenue dans une description suffisamment détaillée de l'autre. Une oeuvre d'art plastique est typiquement matérielle, tandisqu'un poème est typiquement informationnel, en ce sens qu'un exemplaire en vaut un autre. Ainsi un collectionneur n'acceptera pas une copie conforme d'un tableau comme équivalent de ce tableau original, même s'il est incapable de discerner la différence. (Le statut d'un morceau de musique est plus difficile à déterminer.) Par contre, on dit en jurisprudence qu'un billet de banque est fongible, ce qui veut dire que lorsque Pierre rend à Paul « les » cent francs qu'il lui avait prêtés, Paul n'a pas le droit d'exiger que Pierre lui rende le billet même que Pierre lui avait confié.
Nous sommes habitués à traiter les êtres humains comme non fongibles : l'un ne peut remplacer l'autre (même s'ils sont jumeaux). Du point de vue biologique, il est trop tôt pour tâcher de trancher nettement la question de savoir si l'identité d'une être vivant est matérielle ou informationnelle. Toutefois, le rôle de l'information est plus important qu'on n'aurait pu le croire. La dernière façon de caractériser le vivant qui nous occupera dans le présent chapitre repose justement sur une certain notion d'information dont il importe d'esquisser les fondements.
Cette nouvelle notion d'information est complémentaire, à
deux égards, de celle que nous venons d'employer. Premièrement,
elle est susceptible de mesures quantitatives; deuxièmement, elle
fait état de ce que toute information utile est relative à
un certain contexte. Elle n'en demeure pas moins une notion abstraite,
qui reste indépendante de toute considération matérielle.
LA NOTION QUANTITATIVE D'INFORMATION.
Lorsqu'on parle d'informations, on pense d'abord, peut-être, à celles que présentent les journaux ou les bulletins de la TV : il y en a de plus ou moins intéressantes, étonnantes, ou familières. Dégageons deux caractéristiques de cet exemple banal :
(i) L'information est abstraite : elle peut typiquement être transmise par des « media » (mot latin qui signifie « intermédiaires ») très différents les uns des autres.
(ii) L'information n'en est pas une, à moins qu'elle ne nous apprenne quelque chose de nouveau.
Ce sont là des attributs qualitatifs. Mais peut-on mesurer la quantité d'une information? Au premier abord, il peut sembler que cette proposition n'a guère de sens. On a pourtant su lui en donner un. Voyons donc comment.
Partons de l'idée qu'une information est une nouvelle. Si une nouvelle en dit long, c'est qu'elle est surprenante. Pour mesurer l'information, il suffirait donc de savoir mesurer la surprise. Or, on est d'autant plus surpris par un événement qu'il est moins probable. La probabilité, elle, est une quantité, dont la mesure va d'une limite inférieure de zéro (l'impossibilité) à un maximum auquel on donne par convention la mesure 1 (la certitude) (5).
Disons alors, en première approximation, que la quantité d'information que m'apporte une nouvelle donnée équivaut à la mesure de ma surprise. On aurait pu dire que la surprise est simplement l'inverse de la probabilité : plus une nouvelle me paraît probable, moins elle me surprend. Mais il existe une façon plus pratique et intuitivement satisfaisante d'attribuer un chiffre précis à ce degré de surprise. Ce chiffre correspondra au nombre maximum de questions simples dont les réponses permettraient de cerner la nouvelle en question. J'entends par question simple une question à laquelle on peut répondre par oui ou par non. Il s'agit donc de questions qui posent de simples alternatives, ou questions binaires.
Pour mieux comprendre cette idée, imaginons une loterie très simple, dotée d'un million de billets à un franc, numérotés de 1 à 1'000'000, dont un seul est gagnant. A supposer qu'on puisse interroger un oracle qui connaît la réponse, combien de questions binaires faudrait-il pour identifier le numéro gagnant?
La réponse peut surprendre.
On pourrait raisonner ainsi : Puisqu'il y a un million de possibilités, commençons par demander : « Est-ce le 1? », puis « Est-ce le 2? » et ainsi de suite. Le nombre maximum de questions qu'il faudra pour tomber sur la bonne réponse est d'un million moins une; le minimum, une seule. En moyenne, il en faudra donc cinq cent milles. Mais il y a une meilleure méthode, qui met à profit le fait que les nombres sont arrangés en série. Il suffit de trouver un type de question qui permet de réduire chaque fois de moitié le champ des réponses possibles. Le chiffre en question est-il supérieur à 500000? Suivant la réponse, on demande alors : est-il supérieur à 750000 (ou 250000)? et ainsi de suite. On s'aperçoit facilement que selon cette méthode, vingt questions seront suffisantes pour cerner la bonne réponse (6), et on dira alors que la quantité d'information apportée par la nouvelle du numéro gagnant est de vingt unités binaires.
Armés de cette notion quantitative d'information, nous sommes en mesure de présenter la notion d'entropie, qui établira un pont entre la physique et l'information. C'est elle qui sera impliquée dans notre dernière tentative de définition.
Pour donner une assise concrète à l'explication de cette notion, commençons par envisager l'expérience suivante.
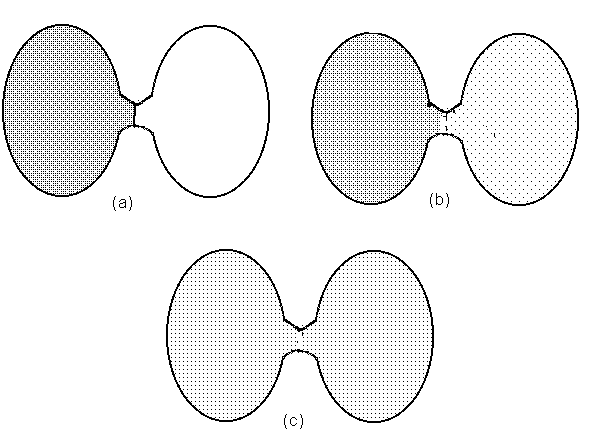
Une paire de vases communicants, symétriques et de grandeur égale, sont séparés par un col pour le moment fermé par une vanne. Dans le vase de gauche, un volume de gaz. Dans le vase de droite, il n'y a aucune molécule quelconque. (Figure 3(a)).
Concentrons maintenant notre attention sur l'instant précis où la vanne s'ouvre. La situation à cette instant se conçoit sous trois aspects : thermodynamique (c'est à dire physique), statistique, et informationnel. Voyons donc ce qu'il en est de chacun de ces points de vue.
(i) (Aspect thermodynamique). Du point de vue de la physique, le vase de gauche exerçait jusqu'à cet instant sur ses parois et sur la vanne une certaine pression. A température constante, selon la loi de Mariotte, la force de cette pression est inversement proportionnelle au volume dans lequel il est contenu (7). D'après la théorie corpusculaire des gaz, cette pression n'est autre que la résultante des impacts des molécules contenues dans ce volume de gaz, dont la trajectoire les amène à se heurter à la paroi. Lorsqu'elles heurtent soit une paroi soit d'autres molécules, les particules rebondissent à la manière de balles élastiques dans une direction déterminée par les circonstances de chaque choc.
Il est à noter que cette pression représente de l'énergie utilisable. (En effet, il suffit de remplacer la vanne par un piston, pour avoir construit l'élément essentiel d'une machine à vapeur.) Mais admettons que nous ouvrons simplement la vanne sans tenter d'exploiter le gradient qui subsiste entre les deux vases. Que va-t-il se passer?
On devine que le gaz se propagera rapidement dans le deuxième vase, jusqu'à ce que chacun des deux vases en contiennent le même volume. Par la même occasion, le gradient de pression entre les deux vases tombera à zéro. La pression moyenne aura baissé . Il sera alors trop tard pour exploiter l'énergie que représentait ce gradient. Cela ne veut pas dire que l'énergie présente dans le gaz est elle aussi tombée à zéro. Si c'était le cas, les molécules individuelles auraient cessé leur danse aléatoire, ce qui reviendrait à dire que leur température serait tombée à zéro. Par conséquent -- et c'est là le point à retenir -- la redistribution du gaz n'a pas éliminé l'énergie inhérente au système, mais elle l'a rendue inutilisable.
Cette inutilisabilité de l'énergie est précisément le sens physique de la notion d'entropie. Le phénomène de diffusion qui vient d'être décrit illustre la deuxième loi de la thermodynamique, qu'on peut formuler ainsi:
Dans tout système clos, l'entropie du système augmentera jusqu'à un maximum; c'est dire que l'énergie utilisable se dégradera inévitablement avec le temps, jusqu'à ce que toute l'énergie contenue dans le système devienne inutilisable.
Physiquement parlant, le système des deux vases communicants a passé, dans l'intervalle des quelques instants qui suivent l'ouverture de la vanne, d'un état d'entropie minimum (représenté par la figure 3(a)) à un état maximum (représenté par 3(c)).
(ii) (Aspect statistique)
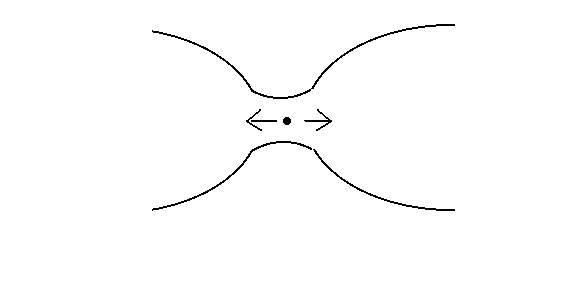
Reprenons maintenant le même phénomène du point de vue de la probabilité. Il n'est plus question sous cet aspect de pressions, de forces ni d'énergie. Le mouvement des molécules de gaz peut être envisagé statistiquement : sans mesurer aucun mouvement individuel, on peut parler de l'énergie cinétique moyenne de toutes les particules qui n'est autre, en physique, que la température du gaz. Rétrécissons notre champ d'observation de manière à ne considérer qu'une seule des molécules qui se trouvent tout près du col qui relie les vases. (Figure 4). Cette molécule, rappelons-le, décrit une trajectoire dont les paramètres individuels ont des valeurs aléatoires. Elle n'a donc pas plus tendance à se diriger vers le haut que vers le bas, ni vers la gauche que vers la droite. Pour simplifier, ne regardons que deux directions possibles : la droite et la gauche. Puisque les deux mouvements sont équiprobables, une molécule placée entre les deux vases se dirigera vers l'intérieur du vase gauche à peu près aussi souvent en moyenne qu'elle se dirigera vers le vase vide de droite. Or, chaque fois qu'une molécule passe à droite, le vide qui caractérisait ce vase devient un peu moins pur. Et jusqu'au moment où il y a plus ou moins le même nombre de molécules à droite qu'à gauche, une molécule quelconque qui se trouve au col aura plus de chances de venir de gauche que de venir de droite, et par conséquent plus de chances de provoquer un changement dans la distribution droite/gauche que de préserver la distribution qui existe jusque là.
Cette simple observation constitue le lien qui unit la perspective statistique aux deux autres. En effet, du point de vue physique, on explique la diffusion du gaz par la pression qu'il exerçait sur la vanne (les anciens disaient que « la nature a horreur du vide »). Cependant, dès que l'on se place au point de vue statistique, horreur du vide ou pression n'apparaissent que comme la conséquence d'un fait purement mathématique, voire une tautologie : si une particule a autant de chances de passer à droite que de rester à gauche, le nombre de particules qui passera de gauche à droite sera proportionnel au nombre de particules qui sont à gauche, et inversement. Donc, tant que les particules de gauche seront plus nombreuses, il y en aura davantage qui passeront à droite.
(iii) (Aspect informationnel)
D'autre part, il est plausible de considérer la phase (a) de la figure 3 comme représentant un maximum d'ordre, et la phase (c) comme représentant un maximum de désordre. Ce qui suggère, si on fait le rapport avec la notion d'entropie, que celle-ci peut être considérée comme une mesure de désordre (8).
L'idée d'ordre et de désordre appartient à l'optique informationnelle que nous avons définie plus haut. En effet, ce qui est hautement ordonné serait un résultat très improbable, donc surprenant, de l'action du hasard; le désordre, au contraire, est conforme à notre attente dans la mesure ou l'on s'attend au plus probable. Or nous venons de voir que la diffusion maximale du gaz dans les deux vases est la simple conséquence de lois statistiques. Donc, puisque l'information représente une sorte d'inversion de la probabilité, l'aspect de notre paire de vases à l'instant où la vanne est ouverte représente une quantité maximale d'information (Figure 3a). C'est là le moment où l'ordre est à son plus haut degré. Petit à petit, l'ordre se désagrège à mesure que l'état du système en question devient plus probable, jusqu'à l'état final de diffusion uniforme qui est dépourvu de contenu informationnel. C'est là précisément l'état d'entropie maximale, qui, nous l'avons vu, correspond du point de vue thermodynamique à l'absence d'énergie utilisable dans le système en question (Figure 3c).
Grâce à ce bref regard sur les notions d'ordre et d'entropie, nous sommes finalement en mesure d'aborder notre dernière définition. Celle-ci peut se résumer dans la formule suivante :
Déf. 6 : Le vivant est ce qui constitue un système dissipatif d'entropie.
Cette formule revient à dire, en langage plus imagé, que la vie représenterait une sorte de rebroussement apparent du cours naturel du monde physique. Voyons pourquoi, en nous plaçant tour à tour à chacun des trois points de vue que nous venons de situer.
(i) Sous l'aspect thermodynamique, le monde vivant présente certaines régions spatio-temporelles du monde où se produisent des « exceptions » locales à la deuxième loi de la thermodynamique. Ces exceptions locales ont été baptisées systèmes dissipatifs d'entropie. Grâce à certaines structures (dont l'origine ultime est encore obscure, puisqu'il s'agit de l'origine de la vie même), certaines concentrations locales de matière acquièrent la faculté de « dissiper » l'entropie qui guette tout système clos. Puisque l'entropie, dans son acception physique, représente une diminution de la quantité utilisable d'énergie, sa dissipation revient à pouvoir tirer de l'environnement une énergie utile, et donc de faire reculer, dans un espace étroitement localisé, la deuxième loi de la thermodynamique. Cette « exception » à une loi de la nature n'est, bien sûr, qu'apparente, puisqu'un système dissipatif n'est justement pas, par définition, un système clos.
Dès qu'une telle capacité de dissipation entropique est quelque peu stable, ses caractéristiques recouperont celles dont nous avons fait l'inventaire plus haut. D'une part, en effet, l'assimilation d'énergie externe et son utilisation au profit du système lui-même traduit assez bien les notions de métabolisme ou d'autopoïèse. Les idées d'intégrité organique et de finalité intrinsèque s'y rattachent aussi de toute évidence, puisqu'on ne pourrait parler d'un système dissipatif, s'il n'y avait pas une frontière relativement bien définie entre ce qui fait et ce qui ne fait pas partie du système. De plus, parler d'utilisation d'énergie présuppose que l'on puisse distinguer ce qui profite au système de ce qui lui nuit.
Seule la reproduction reste apparemment en marge. Pourtant, nous avons vu que si l'on se place au point de vue informationnel, on peut considérer celle-ci comme une stratégie alterne de perpétuation des structures vivantes, qui supplée au métabolisme et à l'occasion (chez les virus) la remplace.
(ii) Sous l'aspect statistique, on peut remarquer que la vie a toujours étonné par son caractère improbable. Cette improbabilité apparente de la vie a conduit plus d'une civilisation à conclure qu'une intention intelligente -- un « créateur » -- devait en être la cause. De manière plus générale, on a tendance à voir dans un événement improbable (une « coïncidence ») un effet de l'intention d'un agent conscient. Nous verrons au chapitre suivant comment la théorie de la sélection naturelle permet de contourner ce raisonnement, qui dans certains contextes est pourtant parfaitement justifiable. Ce qu'il faut retenir pour le moment, c'est que non seulement la probabilité intrinsèque des structures vivantes semble infime, mais encore -- c'est là le sens statistique de l'hypothèse en question -- ces structures ont la capacité de se maintenir dans un état d'équilibre qui semble défier les lois de la probabilité comme elles semblent défier (sous l'aspect physique) la deuxième loi de la thermodynamique. C'est là précisément l'aspect statistique du concept d'un système dissipatif d'entropie.
(iii) Quant à l'aspect informationnel, on a vu qu'il incorpore trois idées. Premièrement, toute structure informationnelle est indépendante de la constitution matérielle qui la réalise. C'est pourquoi, nous l'avons constaté, l'information peut se transmettre à travers une variété illimitée d'intermédiaires ou « media ». Deuxièmement, l'information est quantifiable en vertu de sa relation à la probabilité, ou plutôt à l'improbabilité. Et troisièmement, la notion d'information ainsi quantifiée est étroitement liée à celles d'ordre et de désordre. L'ordre varie donc inversement avec la quantité d'entropie. Notre dernière définition représente donc la vie comme une manifestation locale et temporaire d'un degré d'ordre particulièrement élevé, et qui en vertu de sa propriété « dissipative », a la capacité de maintenir et même d'augmenter cette quantité d'ordre.
Il convient d'insister sur le caractère temporaire et local de ces « rebroussements » de la tendance universelle vers l'entropie que représentent les êtres vivants. Notre dernière définition de la vie implique en effet, comme une conséquence logique, que la vie constitue une exception dans l'univers. Tout système dissipatif a besoin d'un environnement d'où il peut puiser son énergie utile, et dans lequel il peut dissiper son entropie. Dans la mesure où il serait acceptable de considérer l'univers entier comme un système clos (affirmation contestable, d'ailleurs), la vie serait nécessairement vouée à l'extinction ultime. En effet, le système constitué par l'univers entier serait voué par la deuxième loi à la « mort entropique. » Plus modestement, on peut affirmer, contrairement à certaines vues « panpsychistes » qui veulent voir dans l'univers entier une sorte de super-organisme, que la vie est nécessairement tributaire d'un environnement qui n'est pas vivant. Sans lui, ses échanges dissipatifs ne seraient pas possibles. Cette observation est évidemment valable dans la perspective thermodynamique, mais elle ne l'est pas moins lorsqu'on se place aux deux autres points de vue que nous avons relevés. Du point de vue statistique, la vie demeure improbable; du point de vue informationnel, elle représente un degré d'ordre exceptionnel.
De fait, l'isomorphisme qui relie les concepts statistique et physique de l'entropie à des notions informationnelles est si saisissant qu'il nous amène à poser une question radicale : ces trois formules (thermodynamique, statistique, et informationnelle) ne seraient-elles au fond que des aspects d'un seul et unique phénomène? Si la réponse à cette question était affirmative, faudrait-il dire que le niveau le plus fondamental qui sous-tend la nature essentielle de la vie est informationnel?
Les considérations ci-dessus nous donnent deux raisons de le penser, ne fût-ce que dans un esprit spéculatif. Premièrement, nous avons vu que les deux phénomènes qui semblent, sur le plan empirique, être les plus fondamentaux de la vie -- le métabolisme et la reproduction -- peuvent tous deux se comprendre comme des aspects du même processus, à condition de se placer au point de vue informationnel. Deuxièmement, ce qui apparaît comme le corollaire le plus surprenant de la sixième définition, c'est que la seconde loi de la thermodynamique peut se concevoir comme un simple exemplaire d'une tautologie qui tient du calcul des probabilité. Il s'agit, rappelons-le, de la lapalissade que les événements les plus probables arrivent le plus souvent. Or, plus il y a de particules excédentaires d'un côté, plus il est probable qu'elles passent de l'autre. Ainsi, du point de vue informationnel, l'ordre est une exception locale et temporaire à la règle, qui est le Désordre.
CONCLUSION
Notre échantillon de tentatives de définition de la vie nous a permis de voir pourquoi la plupart de ces hypothèses, prises une à une, souffrent de certaines insuffisances : soit qu'elles manquent de clarté, soit qu'elles soient invalidées par des contre-exemples. Cependant chacune d'elles reflète, fût-ce confusément, une intuition digne d'être retenue. En particulier, les notions d'autopoïèse et de reproduction résument assez bien la conception que se fait de la vie le sens commun : la seule considération un peu surprenante que nous ayons hasardée ici est que ces deux caractéristiques ne seraient en fait que deux aspects d'une seule capacité. Cette idée reposait sur la notion d'une perspective informationnelle, qui nous a par ailleurs poussés, dans le cadre de notre sixième définition, vers une exploration sensiblement moins banale.
Il est, certes, permis de douter que notre sixième définition soit, en vérité, une définition à proprement parler. Il ne s'agit pas en tout cas d'une définition de dictionnaire. Toutefois, si l'on la considère comme une définition encyclopédique, il est difficile de nier qu'elle propose une hypothèse assez provocatrice, à savoir que la caractéristique essentielle du monde vivant se situe au niveau de sa structure informationnelle.
Notre analyse, bien qu'inévitablement superficielle, nous invite à poser plusieurs questions difficiles : l'essence de la vie réside-t-elle dans la matière de la vie, ou dans telles structures informationnelles qui la délimitent? Saurait-on concevoir une forme de vie qui ne soit pas organique? Y a-t-il des lois biologiques, si l'on entend par là des lois qui s'appliqueraient à tout être vivant sans exception, que ceux-ci soient terrien, martien, ou issus d'une lointaine galaxie?
Nous aurons l'occasion, dans le cours de ce livre, d'examiner plus avant
ces hypothèse, sous divers aspects. Mais d'abord, et c'est ce qui
nous occupera au chapitre suivant, il convient d'approfondir une question
qui a été explicitement soulevée par la troisième
de nos définitions, mais qui nous a en quelque sorte guettés
tout au long de ce chapitre : qu'est-ce que la finalité, et quel
est son rôle, véritable ou illusoire, dans le monde biologique?
1* Tropismes,
gradients, et circuits cybernétique. On appelle tropisme
les réflexes d'orientation caractéristiques de certaines
plantes, comme le tournesol, ou d'organismes primitifs, tels que les amibes.
Les mécanismes qui gouvernent les tropismes relèvent soit
d'un champ magnétique (géotropisme), soit d'une source
de lumière, (phototropisme), ou enfin d'un gradient chimique,
c'est-à-dire d'une concentration variable d'une certaine substance
(chimiotaxie). Un gradient chimiotaxique fournit une sorte
de téléguidage chimique à tout organisme muni d'un
mécanisme correspondant, qui oriente l'organisme soit dans la direction
d'une concentration de plus en plus forte d'un aliment, soit vers une teneur
de plus en plus faible d'une substance nuisible. Tous ces mécanismes
sont simple en ce sens qu'ils ne tiennent pas compte de leur propre effet.
C'est en quoi ils diffèrent des circuits cybernétiques,
dont la rétroaction ou feed-back leur permet, à chaque
phase d'un comportement, de modifier ce comportement de manière
à s'adapter aux résultats obtenus dans la phase précédente.
Un simple thermostat en fournit un exemple bien connu : dans une première
phase, un thermostat enclenche le chauffage lorsque la température
est inférieure à un certain palier, puis l'éteint
lorsque l'effet de la première phase s'est fait suffisemment sentir.
2. *Comment compter les différents génotypes
possibles à partir d'une seule paire de génotypes mâle
et femelle? Le calcul peut s'esquisser ainsi. L'être humain possède
23 paires de chromosomes. Lors de la reproduction sexuée, la méiose
donne lieu à deux cellules contenant chacune un seul membre de chaque
paire. Ce sont les gamètes, capable de s'unir à une
gamète complémentaire provenant de l'autre parent. Cette
distribution se faisant au hasard, il y a 223 gamètes
possibles chez chaque parent, ce qui donne, pour le génome du zygote
(cellule résultant de l'union des deux gamètes) un total
de 223 multiplié par lui-même, c'est à dire
246 ou environ 1014 (une centaine de trillions) de
combinaisons possibles. Voilà déjà un nombre respectable,
dépassant de loin le nombre des êtres humains ayant jamais
vécu. Cependant, ce chiffre est minuscule comparé au possibilités
réelles. En effet, lorsque les paires de chromosomes se séparent,
leur constitution résulte parfois d'une recombinaison qui modifie
l'identité de chaque membre de la paire. Cela peut se produire (pour
ne citer qu'un seul exemple) lors d'un croisement des deux chromosomes,
qui fait qu'un morceau de l'élément de droite est échangé
contre un morceau correspondant de celui de gauche. De telles fractures
peuvent se faire n'importe où dans chaque paire de chromosomes.
Ainsi, puisque chaque chromosome contient plusieurs milliers de sites de
fracture en puissance, les variantes possibles multiplient encore par un
facteur astronomique le coefficient logarithmique qui dénombre les
éléments distincts qui peuvent constituer les gamètes
possibles. Il en découle un chiffre de zygotes possibles dépassant
facilement les 1065 qu'on estime donner l'ordre de grandeur
du nombre d'atomes dans l'univers.
3. Dans certains cas de rétrovirus tels
que celui auquel on attribue le SIDA, il s'agit plutôt d'un arrangement
caractéristique d'ARN. Rappelons que l'ARN diffère de l'ADN
par l'une des quatre bases dont les combinaisons forment les ?lettres?
individuelles du code génétique.
4.* Mais qu'est-ce, au juste, que la matière?
Une analyse plus poussée de cette notion aura peut-être pour
résultat d'infirmer la distinction même entre matière
et information. En effet, toute matière concrète aura telle
ou telle constitution chimique et physique, à défaut de laquelle
il ne s'agira plus de cette matière-là. Son essence ou sa
nature se cernera donc à partir de certaines propriétés
caractéristiques-- autant dire, de certaines informations.
Dépouillée de ces caractéristiques informationnelles,
il ne s'agira finalement -- comme avait déjà conclu Descartes
dans sa célèbre discussion du morceau de cire qui se transforme
en fondant -- que d'une certaine ?extension?, c'est-à-dire de l'occupation
d'un certain espace dans un certain temps. L'aspect matériel des
choses se réduirait-il donc à telle ou telle région
de l'espace-temps?
5.* Probabilité objective et subjective.
La notion de probabilité désigne un certain formalisme, qui
dérive certains théorèmes à partir d'un petit
nombre d'axiomes élémentaires. Elle désigne aussi
plusieurs interprétations possibles de ce formalisme. Au
premier abord, la notion de probabilité, comme la notion de surprise,
est relative à une situation épistémique (le degré
de ma surprise dépendra de ce que je sais et de ce que j'ignore).
C'est donc une notion subjective. Cependant certaines autres interprétations,
également légitimes, font d'une mesure de probabilité
une quantité objective. Ainsi la probabilité de jeter un
6 en un coup avec un dé juste est objectivement de 1/6 : elle ne
dépend ni de mes croyances, ni de mes connaissances. On verra plus
bas que la notion d'information, qui au premier abord est également
subjective, peut elle aussi représenter des faits objectifs.
6. * Combien de questions? Pour tout problème
dont les solutions possibles peuvent être rangées dans un
ordre fixe, chaque question binaire peut être formulée de
manièe à réduire de moitié le champs des réponses
encore en lisse. Un nombre n de questions suffira donc toujours
à déterminer une solution unique parmi 2n possibles.
7. *La Loi de Mariotte: A température constante,
le volume d'un gaz est inversement proportionnel à la pression qu'il
subit : P x V = Constante.
8. * Ordre, probabilité, information et entropie.
L'ordre est généralement moins probable que le désordre:
voilà pourquoi une table de travail bien en ordre est le résultat
d'un effort sans cesse renouvelé, tandisque le désordre,
lui, semble s'installer spontanément. On peut voir là
une forme informationnelle de la deuxième loi de thermodynamique
qui veut, rappelons-le, que l'entropie augmente d'elle-même
avec le temps. Le rapport avec la notion d'information provient
de la relation déjà notée entre celle-ci et l'inverse
de la probabilité. On peut rendre ce rapport plus intuitif
en soulignant que si l'on prend l'entropie au sens informationnel de ?très
probable?, la seconde loi de la thermodynamique devient une simple tautologie
: la nature tend à produire des états plus probables (de
haute entropie) plus souvent que le contraire. On pourrait donc dire
qu'utiliser de l'énergie, c'est précisément
atteler un appareil à cette tendance mathématique qu'a la
nature de grimper la colline entropique. Par ailleurs, il est a noter que
dans les notions de probabilité et d'information dont
il s'agit ici ont perdu une grande partie de leur aspect subjectif, en
ce sens que l'état entropique d'un système ne dépend
pas des connaissances de tel ou tel observateur. Le lien avec la notion
de surprise n'est cependant pas tout à fait rompu : car un événement
d'une certaine probabilité objective devrait provoquer un degré
correspondant de surprise chez n'importe quel observateur rationnel. Ainsi
nous nous servirons plus loin d'une notion de probabilité intrinsèque
de certaines structures, c'est-à-dire de la probabilité objective
qu'elles fussent le produit d'un jeu de hasard. On s'aperçoit cependant
que la subjectivité n'est pas totalement bannie de cette conception.
En effet, pour la bien comprendre, il faudrait connaître les conditions
initiales dans lesquelles aurait opéré le supposé
jeu du hasard. Or, il est difficile de caractériser objectivement
de telles conditions initiales. A l'intérieur d'un système
dissipatif, par exemple, ce qui serait improbable ailleurs devient hautement
probable.
{kind=link}
{kind=link}
{kind=link}
{kind=link}